![]()
![]()
![]()
![]()
Методология флоуметрии, Âûïóñê 6, 2002 ã. стр.119-129.
Влияние исходного тонуса на динамику нейрогенных констрикторных ответов хвостовой артерии крысы
И.А. Солуянова1, О.С. Тарасова1, А.С. Боровик2, И.М. Родионов1
1Кафедра
физиологии человека и животных Биологического факультета МГУ им. М.В. Ломоносова,
2Лаборатория гравитационной физиологии Учебно-исследовательского
Центра космической биомедицины
Гладкая мышца большинства мелких артериальных сосудов находится в состоянии тонического напряжения [4], величина тонуса определяется балансом констрикторных и дилятаторных факторов, действующих на сосуд. Увеличение или уменьшение тонуса гладкой мышцы ведет к соответствующему изменению просвета сосуда, т.е. изменению его гидравлического сопротивления. Ранее было показано, что изменение исходного тонуса существенно влияет на величину сосудистого ответа при раздражении симпатических нервов [2, 8], однако осталось неясным, изменяются ли при этом динамические характеристики нейрогенных ответов. Разработанная нами методика позволяет изучать нейрогенное сокращение одиночных изолированных сосудов при раздражении нервов, расположенных в стенке исследуемого сосуда, любой заданной последовательностью импульсов электрического тока [1]. Используя специальные паттерны импульсов, можно раздельно исследовать тонический (среднее уменьшение потока) и фазический (изменения сосудистого тонуса, вызываемые ритмически изменяющейся частотой стимуляции) компоненты ответа сосуда [3]. Целью данной работы было исследовано влияние исходного тонуса гладкой мышцы на тоническое и фазическое сокращение сосуда при ритмическом нейрогенном воздействии. Кроме того, мы исследовали, одинаково ли изменяются эти компоненты нейрогенного ответа при создании тонуса с помощью разных констрикторных агентов: норадреналина, серотонина и фенилэфрина.
Материалы и методы исследования
Эксперименты проводили на самцах крыс линии Вистар весом 200-300 г. В качестве объекта исследования была выбрана хвостовая артерия крысы – крупный сосуд мышечного типа.
Экспериментальная установка для перфузии сосудов в режиме постоянного давления была описана ранее [1]. Сегмент сосуда длиной 5-6 мм находился в термостатируемой камере (37° С), заполненной раствором Кребса–Хенселейта, содержащим (мМ): NaCl – 119, NaHCO3 – 25, KCl – 4,7, KH2PO4 – 1,18, MgSO4 – 1,17, ЭДТА – 0,026, D-глюкоза – 5,5, CaCl2 – 2,5. Раствор аэрировали смесью 95%О2 + 5%СО2, pH 7,35. Во время эксперимента измеряли давление на входе и на выходе перфузируемого сосуда, а также объемную скорость тока перфузата. Давление регистрировали с помощью емкостного датчика ДДА-2, для регистрации потока использовали ультразвуковой флоуметр (Transonic System Inc., T106). Проточный датчик (типа 1N) флоуметра встраивали в перфузионную систему перед входной канюлей. Давление на входе (95 мм рт.ст.) и на выходе (75 мм рт.ст.) сосуда создавали гидростатически.
Регистрацию и обработку экспериментальных данных проводили с помощью оригинальной программы на компьютере типа IBM PC с использованием 16–разрядного аналого-цифрового преобразователя (L-card, Москва). Частота оцифровки составляла 10 Гц. Одновременно с регистрацией данных в ходе эксперимента программа управляла работой электростимулятора, задавая последовательность раздражающих импульсов.
Для раздражения периартериальных нервов с обеих сторон сосуда помещали платиновые электроды. Раздражение проводили прямоугольными импульсами тока со сменой полярности, амплитудой 200–300 мА и длительностью 0,2 мс. Ответ сосуда на такую стимуляцию полностью блокировался тетродотоксином (5×10-6 М), т.е. сокращение было нейрогенным.
В начале каждого эксперимента проводили перфузию в течение 30 мин, что приводило к полному расслаблению гладкой мышцы сосуда. Затем сосуд активировали путем трехкратного добавления норадреналина (3×10‑6 М) и последующей стимуляции нервов (20 Гц, 15 с).
Исследовали ответы сосуда при раздражении нервов последовательностью импульсов с заданной средней частотой и мгновенной частотой, изменяющейся по синусоидальному закону (рис. 11.1Г). Такая “синусоидальная” стимуляция имитирует ритмическую активность, наблюдающуюся в эфферентных симпатических нервах в состоянии относительного покоя [6]. Средняя частота следования электрических импульсов в экспериментах была равна 2 Гц, что близко к среднему значению частоты фоновой симпатической активности [4, 6]. Амплитуда модулирующей синусоиды составляла 1 Гц, т.е. частота следования импульсов изменялась по синусоидальному закону от 1 до 3 Гц. Частота модулирующей синусоиды составляла 0,03 Гц, длительность стимуляции – 2 мин, интервалы между воздействиями – 7 мин. Перед “синусоидальной” стимуляцией и сразу после нее в течение 1 мин проводилась стимуляция с постоянной частотой 2 Гц.
Раздражение проводили сначала при исходно расслабленной гладкой мышце, а затем – при перфузии сосуда растворами констрикторных агентов: норадреналина (0,05, 0,1, 0,2, 0,4 и 0,8 мкМ), фенилэфрина (0,015, 0,03, 0,1, 0,3, и 1 мкМ) или серотонина (0,019, 0,038, 0,075, 0,15 и 0,3 мкМ). Для исключения индивидуальных особенностей животных влияние различных констрикторных агентов исследовали на сегментах одного и того же сосуда, чередуя применение веществ для проксимальных и дистальных сегментов. В этих экспериментах определяли: (1) среднее уменьшение потока (тонический компонент сократительного ответа сосуда); (2) амплитуду колебаний потока (фазический компонент сократительного ответа сосуда).
Результаты и обсуждение
При полностью расслабленно гладкой мышце поток через сосуд составлял 4,0-4,3 мл/мин. При раздражении нервов в синусоидальном режиме на фоне тонического сокращения наблюдались колебания потока, частота которых, как и частота модулирующей синусоиды, составляла 0,03 Гц (рис. 11.1.).
Добавление к перфузионному раствору возрастающих концентраций констрикторных агентов приводило к монотонному снижению средней величины потока. При этом изменения тонического и фазического компонентов нейрогенного ответа имели двухфазный характер: при умеренной предактивации гладкой мышцы сосуда они увеличивались, а при более значительной – уменьшались (рис. 11.2.). Максимальные значения этих характеристик нейрогенного сокращения достигались при уменьшении потока через сосуд всего на несколько процентов.
Увеличение тонического компонента было приблизительно одинаковым для всех констрикторных агентов (рис. 11.2А). Это согласуется с результатами наших предыдущих исследований на перфузируемом бассейне задней конечности крысы, свидетельствующих о том, что при создании исходного тонуса с помощью разных констрикторных агентов (в предыдущей работе «веществом сравнения» для норадреналина служил вазопрессин) нейрогенные ответы повышаются приблизительно в равной степени [8]. Вместе с тем следует отметить, что эта зависимость для одиночного сосуда количественно отличается от зависимости для целого сосудистого бассейна. В данной работе максимальное значение тонического компонента наблюдалось уже при незначительном снижении потока, тогда как для системы кровеносных сосудов максимальный ответ соответствовал снижению потока более чем в два раза [8].
Впервые в данной работе было проведено исследование динамических характеристик сократительного ответа сосудов при нейрогенном воздействии. Оказалось, что разные констрикторные агенты (норадреналин, фенилэфрин и серотонин) неодинаково влияют на фазический компонент сократительного ответа. Амплитуда колебаний потока через сосуд, вызванных ритмическим изменением частоты стимулирующих импульсов, при использовании околопороговых концентраций норадреналина была вдвое больше, чем при предсокращении с помощью серотонина или фенилэфрина (рис. 11.2Б).
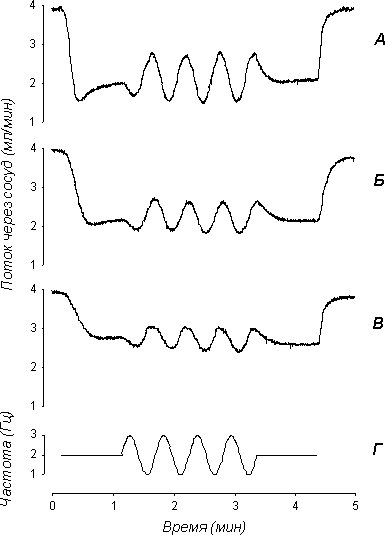
Рисунок 11.1. Изменение потока через сосуд при “синусоидальном” раздражении симпатических нервов. Исходный тонус сосудов создавался с помощью норадреналина (А), серотонина (Б) или фенилэфрина (В). Величина потока при полностью расслабленной гладкой мышце в этих экспериментах была равной 4,1, 4,1 и 4,0 мл/мин, соответственно. На нижнем графике (Г) приведен график изменения частоты электрических импульсов во время стимуляции.
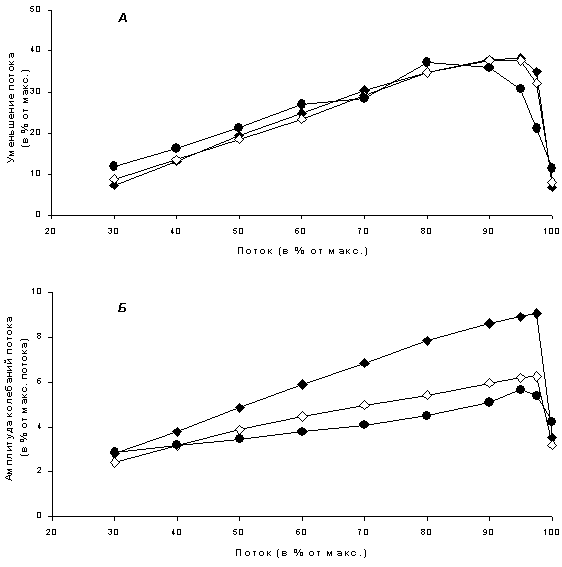
Рисунок 11.2. Изменение тонического (А) и фазического (Б) компонентов нейрогенного вазоконстрикторного ответа при создании исходного тонуса гладкой мышцы с помощью различных констрикторных агентов: норадреналина (черные ромбики, n=5), серотонина (белые ромбики, n=5) или фенилэфрина (черные кружки, n=3). Все параметры (поток перед раздражением нервов, среднее уменьшение потока при стимуляции и амплитуда колебаний потока) нормированы на величину потока через сосуд при перфузии раствором, не содержащим констрикторных агентов. Уменьшение потока соответствует увеличению концентрации вазоконстрикторных веществ
Различное изменение фазического компонента под влиянием норадреналина и фенилэфрина может быть обусловлено их действием на различные типы адренорецепторов в сосудистой стенке: фенилэфрин является селективным агонистом α1-адренорецепторов, а норадреналин также активирует α2-адренорецепторы, расположенные на гладкомышечных клетках, эндотелии и окончаниях симпатических нервов [5]. Поэтому при сокращении сосуда под действием норадреналина в гладкомышечных клетках активируются дополнительные метаболические каскады. Кроме того, экзогенный норадреналин может влиять на секрецию медиатора или на состояние эндотелия, который существенно модулирует ответы сосуда на ритмическую стимуляцию нервов [3].
Известно, что серотонин при действии в низких концентрациях может существенно замедлять расслабление сосуда после сокращения, вызванного норадреналином, причем этот эффект обусловлен его влиянием на механизмы удаления кальция из цитоплазмы гладкомышечных клеток [7]. Такое снижение скорости расслабления может быть причиной меньшей амплитуды фазических колебаний сосудистого тонуса при ритмическом нейрогенном воздействии.
Таким образом, влияние констрикторных агентов на фазический компонент нейрогенного ответа может значительно различаться. Это означает, в частности, что присутствие некоторых вазоактивных веществ в крови может существенно влиять на величину сосудистого ответа на быстрые изменения нейрогенной активности, что скажется на эффективности нейрогенной регуляции сосудистого тонуса.
Исследование выполнено при поддержке Российского Фонда Фундаментальных исследований (гранты № 01-04-48932, 01-04-48475 и 02-04-49193).
Литература
Influence of initial tone on the dynamics of neurogenic constrictor response of rat tail artery
I.A.Soluyanova, O.S.Tarasova, A.S.Borovik, I.M.Rodionov
Department of Human and Animal Physiology, Faculty of Biology, M.V. Lomonosov Moscow State University, Laboratory of Gravitational Physiology, Space Biomedical Center for Training and Research
We studied the response of isolated and isobarically perfused rat tail artery to rhythmic electrical stimulation of intramural sympathetic nerves. The nerves were stimulated by modulated sine pattern (the frequency of stimulation varied by +/- 1Hz around the base value of 2 Hz), the modulating frequency was 0.03 Hz. Sine stimulation evoked frequency-matched oscillations of the flow which were superimposed on the tonic contraction of the vessel. Both mean decrease of flow (tonic component of the response) and amplitude of flow oscillations (phasic component) prominently increased in the presence of constrictor drugs: noradrenaline, serotonin or phenylephrine. An increase of tonic component was similar for all three vasoconstrictor drugs. However, phasic component increased differently, so as during noradrenaline-induced tone it was about two times greater than during preactivation with serotonin or phenylephrine. This indicates that blood content of vasoactive compounds may influence the magnitude of vascular response to fast changes of efferent sympathetic activity.
![]()
![]()
![]()
![]()